Introductory page
I run or better animate a ‘lab’ without benches and without a primary organism or system. The moniker ‘theoretical biology’ sends real biologists who I consort with, fleeing, and the majority of my trainees over the past 2 decades have done experiments in collaborators labs. ‘Modeling’ implies someone hands you data and you deal with it, as best possible, but people in the group did experiments precisely to get the data they needed to theorize about. ‘Data science’ is an oxymoron.
So what are the contours of my subject, call it physical biology, as I conceive it? One can divide the applications of physics to biology as overt or covert. The former would include motor proteins, ion channels, single molecule mechanics, membrane biophysics, cell motility, active matter etc, ie subjects where the physics is manifest. In recent years I have practiced the covert variety, which include models of DNA sequence inspired by statistical mechanics, models of development and cell fate derived from modern (ie post 1960, distinctly not 19th c.) mathematics and the computer science of learning theory. Generally, the motivation is to become more abstract in the hope of attaining more generality. The focus is always on mainstream biological subjects, no bio-inspired physics or niche systems: phenomena first tools second. Beware what I call the ‘hydrogen atom fallicy’. You do not get far in biology by taking a simple system (are there any really?), ignoring the genetics, biochemistry, structure, or whatever pertains, and measuring something (defined by technical feasibility) precisely. There was a period ~20y ago where molecular noise was the rage, in whatever system was at hand. What knowledge can be traced to all those papers, I am at a loss to say. Similarly, one can find data for which a Shannon entropy can be calculated from a snapshot, but the slice of phenomena thereby illuminated is small. A famous paper (Levchenko), in Science of course proved that the information content of a signaling pathway was one bit. It spawned a string of refutations that added to its citation count.
Rather theory is an opportunistic endeavor, like the hedgehog in the essay by Isiah Berlin on Russian novelists, if you want to play the fox, in Berlin’s lexicon, someone wielding an all-encompassing theory, stay with physics. Prime examples of the hedgehog approach in biology would include the Hodgkin-Huxley equations, the Berg-Purcell paper, Delbruck-Luria. All had lasting impact in their respective domains. Other theory has not fared well. Delbruck did not believe the Watson-Crick structure since he reasoned it would be impossible disentangle two cm long polymers of 1nm diameter with 2pi twist per 3nm of length. One strand had to be fragmented. Good fluid mechanics, bad biology, topoisomerases exist. Lord Kelvin definitively inferred the earth was 20-100 million years old from heat flux. One needs to know the limits of the knowable. The hedgehog vs fox dichotomy was expressed with inimitable British wit by Graeme Mitchison in a 2004 article Mitchison_Curr_Bio2004 that has not lost its punch and was accompanied by the photo inspired by those from the Solvay meetings in the early days of quantum mechanics.

From the image, guess who was top dog? It is humbling to read the conference proceedings, since there was more brain power in that group, than any conference I have attended, yet the results did not even anticipate the next decade of biology. Genetic screens defined the terrain.
None of this is to imply that students of theory will not have an impact on biology. A PhD in a theoretical subject plus good taste in biology is very good preparation for deciding what the right experiment to do is. The theorists who worked in partner labs did very interesting experiments (in the judgement of the lab head), that would never have been done by biologists (why? I can only speculate, but the data is solid),
What is a productive level of abstraction? Difficult to define generally but you know it when you see it. Consider the field of ‘synthetic embryology’ or more properly embryo models. (The former term was vetoed by Science magazine as the title of a review on the subject Science_review.) Whatever the moniker, the goal is to construct a viable embryo or organ from embryonic stem cells, following the Feynman mantra of building to understand, which is more relevant to biology than physics. The synthetic route very much facilitates manipulating the pieces to see what causes what, particularly in the case of mouse and human where development is in mother and for human the experiments would contravene the most minimal ethics.
An instance is the micropatterned differentiation of stem cells invented by Warmflash (an ex-theorist), and an image from the paper has been reproduced in at least one development textbook, a more recent sample below.
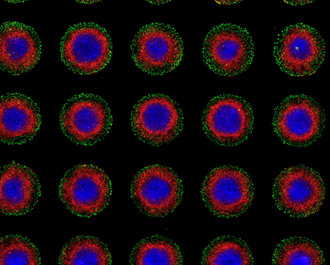
Figure SEQ Figure \* ARABIC 1 Micropattern slide (0.5mm disks) showing reproducibility of patterns after 48h of BMP (thanks to Anna Yoney). Markers: (from outside in) GATA3, BRA, SOX2
To everyone’s surprise, gastrulation (the formation of the body axis and the first step in cell fate specification into ectoderm, mesoderm, and endoderm, the germ-layers) takes place on a coated glass surface in the figure, as best it can. The fetus to be, in a real human embryo, also starts out as an epithelial sheet, more comfortably nestled on a cellular layer of extra-embryonic endoderm. The signaling hierarchy downstream of the first inductive signal is intact (judging by mouse), along with secreted inhibitors, and the topology of the germ-layers arrangement. The synthetic system no doubt butchers the levels, timing, and spatial distribution of the real embryonic signals, but the outcome has many similarities with the embryo showing the canalization present throughout development. The micropatterns are a much more tractable system to study than a 3D embryo implanted in mother, and it’s much easier to knock out genes, fluorescently tag them, etc in cell lines than making a mouse, or contemplate making a monkey. There are now many synthetic embryo models, beginning from the most startling of these by Y. Sasai in 2012, and in my view, they will become a larger component of developmental biology in coming years. Their success will be measured by what they reveal about real embryos.
A well-worn putdown of the synthetic embryologist is the quote of V. Hamburger (a student of Spemann who lived to be a hundred) “Our real teacher has been and still is the embryo—who is, incidentally, the only teacher who is always right”. Hamburger never imagined building an embryo de-novo from cells and achieving a molecular level understanding of self-organization. An analogy should be made with mammalian cell culture (Lasker award to T. Puck in 1958). We learned a great deal of cell biology and signal transduction biochemistry from very aberrant, essentially cancer cells, that could be maintained in culture.